Когда реальность открывает тайны, уходят в тень и меркнут чудеса ...
Пикотехнология белков Издание второе. Исправленное и дополненное.
С представлениями Виктории Соколик можно ознакомиться в первом издании, а во втором издании - с представлениями Александра Кушелева, которые развивались с 1991-го года в т.ч. благодаря авторам, перечисленным в списке литературы.
Глава 3. Аминокислоты и пептидная связь
Введение 97 Кодируемые аминокислоты 98 Пространственная структура аминокислот 104 PDB файлы аминокислот 112 Расчет координат атомов в моделях аминокислот 117 Изомеры кодируемых аминокислот 123 Определение пептидной связи 125 Цис/транс изомерия пептидной связи 127 Ротамерия пептидной связи 130 Заключение 135 Литература к главе 3 136
Введение
Третья глава всецело посвящена пространственной структуре кодируемых аминокислот, которую мы постараемся описать с помощью пикотехнологических моделей. В этом нам
поможет алгоритм расчета координат с учетом
геометрии кольцегранных моделей аминокислот. Программа Молекулярный конструктор
(MC) написана по алгоритму Виктории Соколик. Программы Picotech и Binary_SS написаны по алгоритму Александра Кушелева. Входными данными для программ являются кодирующие белки нуклеотидные последовательности. Выходными данными являются схемы вторичных структур и координаты атомов в стандарте pdb. Файлы PDB для каждой аминокислоты были сопоставлены с
аналогичными файлами координат моделей вьюера PyMol, а также со средними
значениями для случайной выборки экспериментальных данных белков 1icw,
1gs9, 1i3d, 1pjh, 1dds и 2bqz базы данных PDB.
Вторая не менее значимая задача этой главы заключается в описании способа объединения аминокислот посредством пептидной связи. Будет показано три основных пространственных варианта ротамерии: Alpha-, Beta- и Pi-ротамеры, различающиеся между собой поворотом на угол кратный 120 градусов. Модельные эксперименты показали, что повторение Alpha-ротамера пептидной связи между аминокислотными
остатками строит альфа-спираль. Повторение Beta-ротамера – Beta-спираль, Pi-ротамера - Пи-спираль. Случайное чередование ротамеров пептидной связи в полипептидной цепи образует неспиральный фрагмент белка. Некоторые последовательности ротамеров формируют типовые повороты полипептидной цепи белков.
И в заключение данной главы будет приведен геометрический алгоритм
расчёта координат атомов двух смежных аминокислот для одного из ротамера пептидной связи и представлены pdb-файлы для всех ротамеров.
Кодируемые аминокислоты
Аминокислоты являются производными карбоновых кислот, в которых
один из атомов водорода в углеводородном радикале кислоты (как правило, в альфа-положении) замещен на аминогруппу (рис. 3.1). В состав белков входит 19 кодируемых аминокислот (табл. 3.1) и 1 иминокислота пролин (рис. 3.2). Кодируемыми называются аминокислоты, которые кодируются генетическим кодом в составе белков (рис.
3.3).
Таблица генетического кода
Таблица. 3.1. Кодируемые аминокислоты.
Виктория Соколик: Все α-аминокислоты, кроме глицина, имеют асимметрический альфа-углеродный атом (Сα) и существуют в виде двух энантиомеров (L и D). В живых организмах встречаются, как правило, L-аминокислоты (рис. 3.4), поскольку исключительно из этого вида энантиомеров происходит внутриклеточный
синтез белков в рибосоме [1–4]. Это связано с тем, что передача наследственной информации возможна только в хирально чистой среде, а также с хиральной стабильностью аминокислот. D-аминокислоты в составе белков или пептидов возникают крайне редко и только посттрансляционно [5], вследствие самопроизвольной или ферментобусловленной изомеризации отдельных аминокислотных остатков. Под хиральной молекулой понимают молекулу с несимметричной структурой, которая не может быть наложена на ее зеркальный образ
путем трансляции или вращения. Тогда как энантиомер – это изомер хиральной молекулы с хиральностью определенного типа. В этом смысле кодируемые аминокислоты являются гомохиральными. Гидроксипролин, треонин,
изолейцин имеют два асимметричных атома и образуют по две пары энантиомеров.
Лево- и правосторонние хиральные молекулы долгое время считались
энергетически эквивалентными, точными зеркальными отражениями друг друга. Как же так? Зеркально не совпадают, а энергетически совпадают?
В 1957 году физиками было сделано удивительное открытие: оказалось,
что, например, при бета-распаде электроны с право- и лево-ориентированными
спинами рождаются с разной частотой, то есть нарушается зеркальная симметрия (иначе – "несохранение четности") [6]. Казалось бы, этот эффект должен
проявляться только на электронном уровне. Но созданная А. Саламом и
С. Вайнбергом объединенная теория слабого и электромагнитного взаимодействия предсказала (и это подтвердили экспериментально), что "электрослабые"
силы, действующие между электронами и атомным ядром – его протонами и
нейтронами, тоже будут отличать правое от левого [7]. Поэтому атомы и молекулы, которые прежде рассматривали как зеркально симметричные, обретают
101
хиральность, а оптические изомеры становятся энергетически неодинаковыми –
возникает обусловленный нарушением четности энергетический сдвиг (НЧЭС).
Квантовомеханические расчеты показали, что L-аминокислоты имеют меньшую энергию, а статистическая физика утверждает: в любой равновесной системе объекты с меньшей энергией должны встречаться чаще. И чем значительней будет НЧЭС двух энантиомеров, тем сильнее при синтезе из простых
веществ будут различаться их концентрации. Для аминокислот разница концентрации составляет 10-17. Поэтому, если предположить, что это начальная бесконечно малая асимметрия ответственна за доминирование L-изомеров, то она
должна многократно усиливаться в результате биохимических эволюционных
процессов. Предложено несколько механизмов усиления, в том числе "бифуркационных катастроф" Кондепуди [8]. Он включает автокатализ и антагонизм
энантиомеров. При определенных условиях один вид в конечном итоге полностью вытесняет другой. Но для этого необходимо, чтобы разница концентрации
была 10-17, в противном случае тепловые флуктуации подавляют эффект усиления. С единой изомерией протеиногенных аминокислот связывают различия
живого и неживого вещества, в то время как белки абиогенного происхождения
являются рацематами, поскольку содержат примерно равные количества молекул любой из двух (L или D) возможных пространственных структур. Хиральная чистота живых организмов принципиально важна: полимеры хирально нечистого состава менее прочны, медленнее растут и быстрее разрушаются, чем
хирально чистые.
В зависимости от состава и структуры радикала аминокислоты характеризуются специфическими свойствами (табл. 3.2). К алифатическим аминокислотам относятся глицин, аланин, валин, лейцин и изолейцин. Эти аминокислоты не несут в боковой цепи гетероатомов (N, О или S), циклических группировок и характеризуются отчетливо выраженной низкой полярностью.Также
малополярны серосодержащие аминокислоты – метионин и цистеин, причем
цистеин существует лишь в недиссоциированном состоянии. Благодаря образованию дисульфидных мостиков, цистеин выполняет важную функцию стабилизации пространственной структуры белков. Аминокислота цистин состоит из
102
двух остатков цистеина, соединенных дисульфидным мостиком. Ароматические аминокислоты содержат циклы. В этой группе лишь фенилаланин проявляет низкую полярность. Тирозин и триптофан характеризуются заметной, а гистидин – даже высокой полярностью.
Александр Кушелев:
Так изображают аминокислоты в "современных" учебниках молекулярной биологии...
Рис.1
В настоящее время считается, что существует две формы аминокислот, L-левая и D-правая. В действительности существует четыре пространственных изомера аминокислот.
Рис.2
Если поменять местами радикал и водород, присоединенные к тетраэдрическому атому углерода, то L1-форма поменяется на L2-форму или наоборот. Однако построенная из аминокислотных остатков спираль не поменяется с правой на левую или наоборот. В чём же дело? А дело в том, что перестановка радикала в позицию водорода не меняет положение атома азота и, следовательно, ход основной цепи. Это значит, что существует не 2, а 4 формы (пространственных изомера) аминокислот: L1, L2, D1, D2. Перестановка радикала меняет форму L1 на L2 или D1 на D2, и это не влияет на ход основной цепи. Если же поменять форму L1 на D1 или D2 на L2, т.е. поменять местами водород и азот, то радикал останется на прежнем месте, а ход основной цепи изменится, т.е. превратится в своё зеркальное отражение. Именно такое изменение изомера и показано на рис.2 Обратите внимание на то, что изомеры L1 и L2 не являются зеркально симметричными. Это же относится и к изомерам D1 и D2
Аминокислота глицин в качестве радикала имеет водород, поэтому глицин действительно имеет лишь две формы L и D. Другими словами L1 и L2 можно отличить лишь заменив один из атомов водорода на другой изотоп. В этом случае L1 и L2 изомеры глицина будут иметь разные моменты инерции. При этом L1 и D1 изомеры будут зеркально симметричными, но иметь одинаковые моменты инерции.
Как же выяснить, какая форма аминокислот используется в белках, L1 или L2 ? Это легко выяснить на моделях циклических фрагментов белков, например, лизоцима:
Цитата: Дисульфидный мостик замыкает цикл лизоцима только в том случае, если фрагмент собран из L1-пространственных изомеров, т.е. таких, у которых атом CB1 лежит в плоскости пептидной группы. Если же радикал поменять с водородом, то дисульфидный мостик разомкнётся, что противоречит результату химического эксперимента. Таким образом, с помощью модельного эксперимента строго доказано, что кодируемые аминокислоты являются L1-пространственными изомерами, т.е. L-изомерами, у которых CB1-атомы радикалов лежат в плоскости пептидной группы. В процессе этого исследования были сделаны очередные научные открытия.
1
- чем ниже, тем менее вероятна мутация соответствующей аминокислоты. 2
- гидрофобность - произвольные единицы основанные на OHM шкале, которые выражают
возможность замены одной аминокислоты другой в ходе эволюции [9]. 3
- вероятность того, что >5% поверхности аминокислоты соприкасается с раствором, окружающим белок.
Нейтральные аминокислоты содержат гидроксильные (серин, треонин)
или карбоксамидные группы (аспарагин, глутамин). Молекулы аспарагина и
глутамина высоко полярны. Карбоксильные группы боковых цепей кислых
аминокислот – аспарагиновой и глутаминовой – полностью ионизированы во
всем диапазоне физиологических значений рН. Аналогичным образом боковые
цепи основных аминокислот – лизина и аргинина – полностью протонированы
в нейтральной области рН. Сильно основной, а, следовательно, очень полярной аминокислотой, является аргинин, содержащий гуанидиновую группировку.
Особое положение занимает пролин. Пролин, строго говоря, является не амино-
, а иминокислотой (рис. 3.2). Атом азота в кольце является слабым основанием
и не протонируется при физиологических значениях рН. Благодаря циклической структуре пролин вызывает изгибы полипептидной цепи, что очень существенно для структуры белков.
Гидрофобность аминокислот выражает степень, в которой они предпочитают неполярное окружение (этанол в качестве растворителя или внутренность
белка) по сравнению с полярным окружением, таким как вода (рис. 3.5). На
этой диаграмме гидрофобные аминокислоты располагаются ниже нуля, а гидрофильные – выше. Используются две шкалы. Шкала Фроммеля (Frommel,
1984) выражает свободную энергию переноса из гидрофобной среды в воду.
Это внутреннее свойство аминокислоты, не зависящее от роли аминокислоты в
белке. OMH шкала (Sweet and Eisenberg, 1983) – мера того, насколько вероятна
замена данной аминокислоты другой гидрофобной аминокислотой [9, 10].
Пространственная структура аминокислот
Виктория Соколик: В настоящее время для изображения пространственной структуры аминокислот наибольшее распространение получили шарико-стержневые модели и
их варианты (рис. 3.6), в которых межцентровые расстояния и торсионные углы
между химическими связями рассчитаны по квантово-механическим функциям.
Рис. 3.6. Разнообразие шарико-стержневых моделей аминокислот.
В наших моделях аминокислот для визуализации атомов и их групп мы
использовали преимущественно «огранённые шарики» – октаэдры, которые в
отдельных случаях моделировали из колец-электронов, что существенно увеличивало разрешающую способность модели. Это обусловлено тем, что достраивание до завершения внешних электронных оболочек в группах атомов NH2,
OH, CO, СαН и R происходит до 8 колец-электронов, т.е. до октаэдра. Поэтому
при построении данных моделей были заданы только атомные радиусы (табл.
2.2) для атомов С, О, N, Н и S, а торсионные углы автоматически устанавливались при совмещении граней соответствующих октаэдров. Таким образом,
шаблоном аминокислот может служить модель, представленная на рисунке 3.7,
где R–радикал, уникальный для каждой аминокислоты.
Рис. 3.7. Пикотехнологически модели аминокислот.
А – многогранный, В – кольцегранный, С – кольцегранный с цветной маркировкой спинов электронов молекулярной электронной оболочки.
А О В С
С
Сα
NH2
OH
R
NH2
OH
О
С
R
Сα
105
Мы моделировали шаблон L-энантиомера аминокислот, в котором асимметричный Сα атом характеризуется sp3
-гибридизацией электронов и тетраэдрической симметрией заместителей.
Проанализируем структуру радикалов аминокислот (табл. 3.3). Во 2-м и
3-м стобцах таблицы представлены смоделированные из октаэдров аминокислотные остатки со специфическими радикалами. Их структурное сходство с используемыми моделями в молекулярной биологии (1-й столбец таблицы) свидетельствует об адекватности нашего методологического подхода.
Таблица. 3.4. Кольцегранные модели аминокислот А.Ю. Кушелева
PDB файлы аминокислот
В качестве pdb. файла любой аминокислоты или белка используют запись
координат каждого атома структуры в декартовой системе координат в общепринятом формате. В таблице 3.5 представлен пример формата pdb. файла для
небольшого участка белка.
В этой главе мы проведём сравнительный анализ pdb. файлов моделей
аминокислотных остатков, построенных нами, с аналогичными
моделями, которыми оснащены современные вьюеры, и рассчитанных современными методами молекулярной динамики по экспериментальным данным
рентгеноструктурного анализа (РСА) или ядерного магнитного резонанса
(ЯМР). В таблице 3.6 приведены координаты [x, y, z] для атомов 20 кодируемых аминокислот, приведенных в единое исходное положение пространства.
Во 2-ом столбце таблицы представлены pdb. файлы моделей аминокислотных
остатков, вычисленные в авторской программе Molecular
Constructor, в 3-ем и 4-ом – рассчитанные по квантово-механическим функциям
113
для вьюеров или полученные в результате конформационного анализа данных
«мокрого» эксперимента.
Заметим, что модели Соколик отличаются от моделей Кушелева только положением
радикала в аминокислоте. В расчёте основной цепи белковой молекулы различий нет. В настоящее время алгоритм позволяет точно определить координаты атомов во всех спиральных участках, т.е. альфа, бета, пи, 310. Что касается изломов на остатках Met и Pro, то эта часть алгоритма ещё не реализована. Таким образом, последняя версия программы (Binary_SS, ноябрь 2024г) гарантирует правильность только 2D схемы любого белка, который строится рибосомой по коду. Пространственные модели и соответственно PDB файлы могут требовать проверки и доработки в ручном режиме. Тем не менее, лучше иметь правильную схему вторичной структуры белка, чем условную пространственную структуру, где неизвестно, из каких спиралей (альфа- или пи-) построен белок. РСА и ЯМР не отличают правую альфа-спираль от левой пи-спирали, поэтому реальная погрешность этих методов превышает 5 аминокислотных остатков вдоль первичной последовательности белка. Подробности можно будет прочесть в статье: "3D Genetic Code", которая готовится к публикации в настоящее время.
Изомеры аминокислот
Из кодируемых аминокислот серин, лейцин и аргинин имеют структурные
изомеры, которые кодируются различными кодонами. Структура и координаты
атомов изомеров этих трёх аминокислот уточняются. Остальные аминокислоты представлены в белках только в единственном
изомерном варианте.
Определение пептидной связи
Пептидная связь – вид амидной связи, возникающей при образовании
пептидов и белков в результате взаимодействия α-аминогруппы (—NH2) одной
аминокислоты с α-карбоксильной группой (—СООН) другой аминокислоты.
Пептидная группа -С(О)-NH- в белках находится в состоянии кето-енольной
таутомерии (рис. 3.12).
Рис. 3.12. Резонансные формы пептидной группы.
Виктория Соколик: Благодаря таутомерии пептидная связь имеет на 40% характер двойной
(сопряжённой) связи, что проявляется в уменьшении её длины (на 10%) по
сравнению с длиной одинарной -С-N- связи. Исходя из найденного значения
длины пептидной связи (1,33 нм), Pauling L. и Corey R. представили простую
резонансную модель, в которой пептидная связь находится в заряженной форме
чуть больше 1/3 всего времени [12]. Иными словами, распределение электронов
внутри пептидной группы характеризуется смещением электронного облака к
атому кислорода (О), в результате чего атом азота (N) приобретает частичный
положительный заряд.
Сопряжённый характер пептидной связи обусловливает планарность пептидной группы (все Cα
i, C’i, Oi, Ni+1, Hi+1 и Сα
i+1 атомы находятся в одной плоскости). На рисунке 3.13 представлены плоскости пептидной связи в полипептиде. Небольшие отклонения от планарности (Δω < 20о
) считаются энергетически
приемлемыми.
126
В 1888 г. Данилевский А.Я. в своей «теории элементарных рядов» впервые высказал идею -C-N- связи между остатками аминокислот в белковой молекуле [13]. В 1900 году немецкий химик-органик Fischer E. [14] выдвинул пептидную теорию строения белка. И уже в 1902 году он получил неопровержимые доказательства существования пептидной связи, а к 1905 году разработал
общий метод, при помощи которого стало возможным синтезировать пептиды в
лабораторных условиях [15, 16].
Особенности пептидной связи:
- возможность существовать в двух резонансных формах: кето- и енольной;
- является на 40% двойной, что проявляется в уменьшении её длины до 1,33 Å
вместо 1,47 Å и сильно заторможенному вращению групп атомов по оси данной
связи;
- копланарность структуры – все атомы, входящие в пептидную группу, лежат в
одной плоскости (рис. 3.13);
- способность к образованию водородных связей, причем каждая пептидная
группа может формировать две водородные связи с другими группами, в том
числе и пептидными (кроме образованных пролином и гидроксипролином), что
играет важную роль в сворачивании полипептидной цепи.
Александр Кушелев: Пикотехнологическая модель аминокислоты показывает, что всё гораздо проще.
Проблема 3D моделирования процесса сборки белковой молекулы заключается в том, что кодируется только угол фи, а меняются "по месту" ещё два угла.
Кодируемый угол фи. Красный шарик показывает альфа-ротамер, синий - пи-ротамер, а зелёный - бета-ротамер.
Второй (транспозиционный) угол (тета) не кодируется. Он принимает разные значения в альфа-, бета-, пи-, 310-спиралях. Его величину определяет длина водородной связи в соответствующей спирали.
В третьей главе представлены модели протеиногенных аминокислот, построенные по следующим принципам:
- структура аминокислот является трёхмерной;
- атомы в моделях аминокислот могут быть аппроксимированы правильными многогранниками;
- многогранные модели аминокислот дают возможность рассчитать координаты центров атомов в декартовой системе с помощью геометрического алгоритма вручную или в программе;
- межатомные расстояния определяются ковалентными радиусами атомов,
тогда как углы между связями детерминируют грани многогранников;
- структурный шаблон атомов, общих для всех аминокислот, описывается
одинаковыми координатами.
Сравнительный анализ матрицы координат, рассчитанной геометрическим способом для многогранных моделей аминокислот, с соответствующими экспериментальными данными показал соответствие моделей результатам "мокрых экспериментов". Кроме самих аминокислот в этой главе мы уточнили способ объединения
аминокислот в полипептидах, а именно рассмотрели три вида ротамеров пептидной связи. В модельных экспериментах было выявлено, что повторение альфа-, бета-, пи-, 310-ротамеров пептидной связи обусловливает формирование соответствующих спиралей. Механизм трансляции мы рассмотрим в следующей главе.
Литература к главе 3
1. Flack H.D. Louis Pasteur’s discovery of molecular chirality and spontaneous
resolutionin 1848, together with a complete review of his crystallographic and
chemical work // Acta Crystallography. – 2009. – A65. – P.371–389.
2. Lamzin V.S., Dauter Z., Wilson K.S. How nature deals with stereoisomers//
Curr. Opin. Struct. Biol. – 1995. – 5. – P. 830–836.
3. Mason S. F. Origins of biomolecular handedness //Nature. – 1984. – 311. – P.
19–23.
4. Prelog V. Chirality in chemistry //Science. – 1976. – 193. – P. 17–24.
5. Lam H., Oh D.C., Cava F., Takacs C.N., Clardy J., de Pedro M.A., Waldor
M.K. D-amino acids govern stationary phase cell wall remodeling in bacteria
//Science. – 2009. – 325. – P. 1552–1555.
7. Вайнберг С. Мечты об окончательной теории // Dreams of a Final Theory.
– М.: ЛКИ, 2008.– С. 256.
8. Kondepudi D.K. Selection of molecular chirality by extremely weak chiral interactions under far-from-equilibrium conditions //Biosystems. – 1987. – 20
(1). – P. 75-83.
9. Sweet, R.M. and Eisenberg, D. Correlation of sequence hydrophobicities
measures similarity in three-dimensional protein structure // J. Mol. Biol. –
1983. – 171. – P. 479-488.
10.Frommel C. The apolar surface area of amino acids and its empirical correlation with hydrophobic free energy // J. Theor. Biol. – 1984. – 111. – P. 247-
260.
11.Соколик В.В. Геометрия аминокислот / Материалы XV Международной
научно-практической конференции «Наука и современность – 2012»,
14.03.2012, Новосибирск. – С 13-18.
12.Pauling L., Corey R., Branson H. The structure of proteins: Two hydrogenbonded helical configurations of the polypeptide chains // Proc. Natl. Acad.
Sci. USA. – 1951. – 37. – P. 205-211.
137
13.Данилевский А.Я.Отношение анатомической и химической дифференцировки главных белковых веществ мышечной ткани к характеру ее деятельности // Физиологический сборник. І. – Харьков, 1888. – С. 331–346.
14.Fischer E. Untersuchungen über Aminosaüren, Polypeptide und Proteine // Ber
Deutsch Chem Ges. – 1906. – 39. – P. 530-610.
15.Каліман П.А. Харківський національний університет ім. В.Н. Каразіна
біля витоків вітчизняної біохімії // Укр. біохім. журн. – 2007. – 79, № 1. –
С. 137-147.
16.Шамин А.Н. История химии белка. Серия «Из наследия А.Н. Шамина»,
Изд. 2, 2006. – 233 с.
17.Лалаянц И.Э., Яковенко Л.В. Роль стереоизомерии в биохимической регуляции // Биология. Прил. к газете Первое сентября. - 2009. – № 17. – С.
2-5.
18.International Union of Pure and Applied Chemistry, Organic Chemistry Division, Commission on Nomenclature of Organic Chemistry, Nomenclature of
Organic Chemistry, Sections A, B, C, D, E, F, and H, 1979 Edition, J. Rigaudy
and S. Klesney, eds, Pergamon Press, Oxford, 1979. – 559 p.
19.Gal J. Louis Pasteur, language and molecular chirality.I. Background and Dissymmetry //Chirality. – 2011. – 23. – P. 1−16.
20. Flack H.D. Louis Pasteur’s discovery of molecular chirality and spontaneous
resolution in 1848, together with a complete review of his crystallographic and
chemical work // Acta Cryst. – 2009. – A65. – P. 371–389.
21.Твердислов В.А., Яковенко Л.В., Жаворонков А.А. Хиральность как проблема биохимической физики // Журнал Рос. хим. об-ва им. Д.И. Менделеева. – 2007. – LI, №1. – С. 13-23.
22.Pauling L. The nature of the chemical bond, 3rd. ed., Cornell University Press,
1960. – 644 p.
23.Pauling L., Corey R.B. Atomic coordinates and structure factors for two helical
configurations of polypeptide chains// Proc. Natl. Acad. Sci. USA. – 1951. –
37. – P. 235−240.
138
24.Jabs A., Weiss M. S., Hilgenfeld R. Nonproline cis peptide bonds in proteins //
J. Mol. Biol. – 1999. – 286. – P. 291-304.
25.Лалаянц И.Э. Хиральность на потоке / по материалам New Scientist. –
2002. – № 2329. – Р. 19. http://him.1september.ru/2004/28/47.htm.
26. Lummis S.C., Beene D.L., Lee L.W., Lester H.A., Broadhurst R.W.,
Dougherty D.A.Cis-trans isomerization at a proline opens the pore of a neurotransmitter-gated ion channel // Nature. – 2005. – 438 (7065). – P. 248-252.
27.Соколик В.В. Способ моделирования пространственной структуры белка
по детерминирующей его нуклеотидной последовательности // Биофизический вестник. – 2010. – Вып. 24 (1). – С. 31-45.
28. Соколик В.В. Моделирование пространственной структуры белка по детерминирующей его нуклеотидной последовательности / Материалы VI
Международной научно-технической конференции «Актуальне вопросытеоретической биофизики, физики и химии». Тезисы докладов БФФХ2010 (26-30 апреля 2010 г.), T.1, Севастополь.– С. 201-204.
29.Sokolik V.V. Modeling of the individual structural template of protein on
determining it nucleotide sequences / Материалы VII Международной конференции по биоинформатике, регуляции и структуры геномов и системной биологии (BGRS\SB-2010). Тезисы докладов (20-27 июня 2010 г.),
Новосибирск. – С. 275.
30. Соколик В.В. Пространственная структура гомологов основного актина и
α-актина 1 различна // Сборник материалов I Международной научнопрактической конференции «Наука и современность – 2010» в 3-х частях
/ Под общ. ред. С.С. Чернова - Новосибирск: «СИБПРИНТ», 2010. – 278
с. Ч. 1, С. 41-46.
31.Соколик В.В. Ротамерные варианты конфигурации пептидной связи и их
кодирование в геноме / Материалы X Украинского биохимического съезда. Тезисы докладов (13-17 сентября 2010 г.), Одесса. – С. 105-106.
32.Кушелев А.Ю., Полищук С.Е., Неделько Е.В. и др. Построение масштабной модели структуры белка // Актуальные проблемы современной
науки. – 2002. – №2. – С. 236-243.
139
33.SokolikV.V. Algorithm of protein structural template decoding according to its
determined nucleotide sequence / Fist International Conference “Fundamental
Medicine: From Scalpel toward Genome, Proteome and Lipidome” (April 25-
28, 2011), Pax Grid Virtual Conferences, Kazan, Russia. – P. 117-119.
34.Соколик В.В. Карта Рамачандрана: ротамерия пептидной связи и фолдинг
белка / Материалы VII Международной научно-технической конференции «Актуальне вопросы теоретической биофизики, физики и химии».
Тезисы докладов БФФХ-2011 (26-30 апреля 2011 г.), Севастополь.–
С.137-139.
35.Sokolik V.V. Protein is coded in genome and synthesized in ribosomes as a
structural template of a rotameric version sequence of peptide bound configuration / The International Moscow Conference on Computational Molecular
Biology, МССМВ-11 (July 21-24, 2011), Moscow, Russia. – P. 347-348.
36.Кушелев А.Ю., Соколик В.В. Пикотехнология – новый подход в моделировании пространственной структуры белка // Заочная Международная
научно-практическая конференция «Современная наука: тенденции развития», Краснодар: НИЦ Априори. – 2012. – С. 203-207.
Приглашение к сотрудничеству
На базе научного открытия нами создан онлайн-сервис по определению структуры белковых молекул. Теперь мы сможем зарабатывать вместе.
По старой технологии определение одной структуры белка обходится примерно в 10 000 евро, а ждать нужно от 2 месяцев до 3 лет. По новой технологии структура определяется в 1000 раз точнее и в миллиард раз быстрее. 80% от найденного Вами заказа принадлежат Вам, как менеджеру.
Наш лозунг: "В 1000 раз лучше, в 1000^3 быстрее и в 1000 раз дешевле!"
Ваша задача заключается в размещении рекламы на онлайн-сервис белковых структур. Рынок этих структур очень большой и продолжает стремительно расти. Ежедневно кто-то оплачивает до 60 структур по средней цене 10 000 евро за штуку. Новая технология позволила на одном персональном компьютере за неделю определить структуры всех 115 000 белков человека, для которых известна нуклеотидная кодирующая последовательность. При этом качество результата, полученного по новой технологии в 1000 раз выше по точности, в миллиард раз по быстродействию и в 30 раз шире по номенклатуре белковых молекул. Единственное, что нам сегодня не хватает - рекламы.
Как получить Вашу первую зарплату менеджера? Найти заказчика белковых структур и убедить его заказать за счёт лаборатории Наномир пробный заказ. Когда заказчик распробует новую технологию, он начнёт делать коммерческие заказы. С первого коммерческого заказа менеджер получает 80%. С последующих заказов процент будет постепенно уменьшаться, но с первого заказа другого заказчика менеджер снова получит 80%. Зарплата менеджера может достичь миллиона евро в день. И это не предел.
Инвестирование научных проектов
Приглашаем инвесторов и меценатов.
Как продвинуть цивилизацию на новый уровень своего развития и получить при этом огромные прибыли?
- Вложить деньги в научные разработки.
Новейшие виды экологически чистых и мощных источников энергии, средство для продления жизни, высокие технологии.
Все это реально создать в ближайший год-два при наличии достаточного финансирования.
Готовые коммерческие продукты
1. Online service PROTEIN PICOTECHNOLOGY
2. Сверхдобротные одномодовые диэлектрические резонаторы в т.ч. с большим диапазоном перестройки
3. Станки для производства высокодобротных одномодовых резонаторов
4. Технология изготовления сапфировых линз
5. Магнитный тороидально-сферический конструктор
Проекты
01 Ruby Emdrive (Микроволновый двигатель без реактивной струи)
02 Ruby Power Source (Микроволновый источник энергии)
03 Средство продления жизни (Возвращение молодости)
04 Октаэдрический редуктор
05 Шестеренчатая передача Кушелева
06 Магнитный подвес-стыковка-герметизация модулей
07 Ионно-микроволновый фрактальный излучатель
08 Гибкий отражатель из жестких элементов
09 Энциклопедия "Наномир"
10 Экспертиза
11 Конструктивные компьютерные игры
12 Интеллектуальный кодовый замок
13 Очки кругового обзора
14 Тетраэдрический сканер
15 Программируемая архитектура
16 Источник энергии промышленной частоты
17 Источник энергии постоянного тока
18 Монокристаллическая видеокамера
19 Система определения активных участков белка
20 Тераваттный лазер непрерывного действия
21 Бактериальный синтез алмазов
22 Шестеренчатые передачи с тремя степенями свободы
23 Сверхсветовая связь
24 Безосевая шестеренчатая передача
25 Aктивный язык программирования
26 Телевидение миллиметрового и оптического диапазонов
27 Микроволновая архитектура
28 Компьютерный экран из автономных элементов
29 Чтение / запись ДНК
30 Сверхсветовая локация / зрение
31 Нейтрализатор акустического сигнала
Коммерческое предложение:
Виктория Соколик: Уважаемые коллеги, Вашему вниманию предоставляется услуга -- моделирование 2D и 3D структуры любого белка без ограничений в его размере и степени изученности с помощью программного обеспечения, базирующемся на принципиально новом подходе декодирования нуклеотидной последовательности, детерминирующей данный белок.
Всё, что необходимо от заказчика, это нуклеотидная последовательность мРНК интересующего его белка (или код этой нуклеотидной последовательности в EMBL, или хотя бы код самого белка в PDB).
В течение 1-3 суток мы готовы предоставить Вам схему вторичной структуры заказанного белка (2D), модель его пространственной структуры (3D) в виртуальном пространстве, а также файл .pdb с координатами каждого атома белка.
Файл .pdb может быть использован по аналогии с файлами закристаллизованных белков из PDB банка для дальнейшего конформационного анализа белка методами молекулярной динамики с учётом физико-химической специфики микроокружения белка или его взаимодействия с лигандами.
Таким образом, Вы сможете максимально быстро удобным для Вас способом (по электронной почте, на сайте либо на электронном носителе) получить информацию о структуре Вашего белка.
Сотрудничество может быть различным:
- участие в научных дискуссиях на форуме (конструктивное)
- совместное создание коммерческого продукта
- поиск инвесторов
- выступить менеджером по продаже готовых коммерческих продуктов
- конструктивные предложения по продвижению идей лаборатории Наномир
- содействие в проведении экспериментов и т.п.
- написание совместных научных статей и т.п.
- материальный вклад (денежный или обеспечение оборудованием и материалами)
Пожалуйста, сообщайте о своем вкладе, чтобы мы зачли Вас как партнера лаборатории Наномир.


















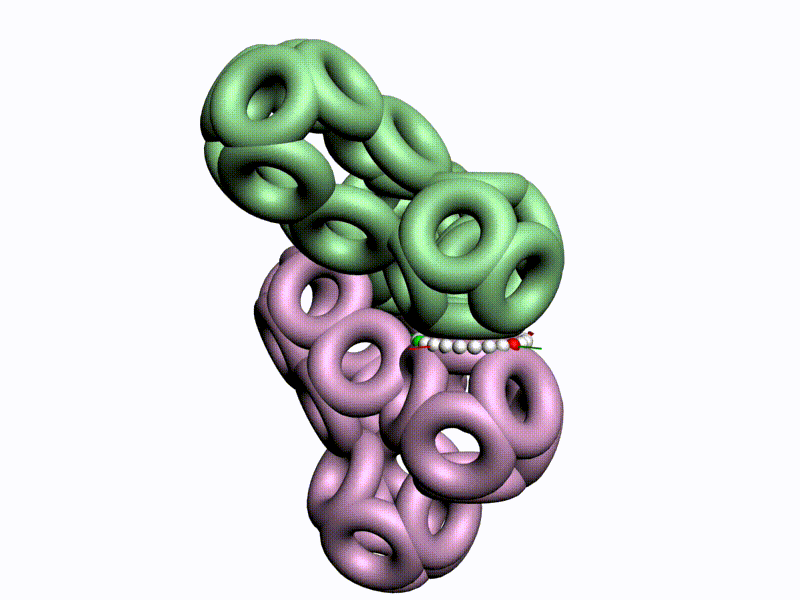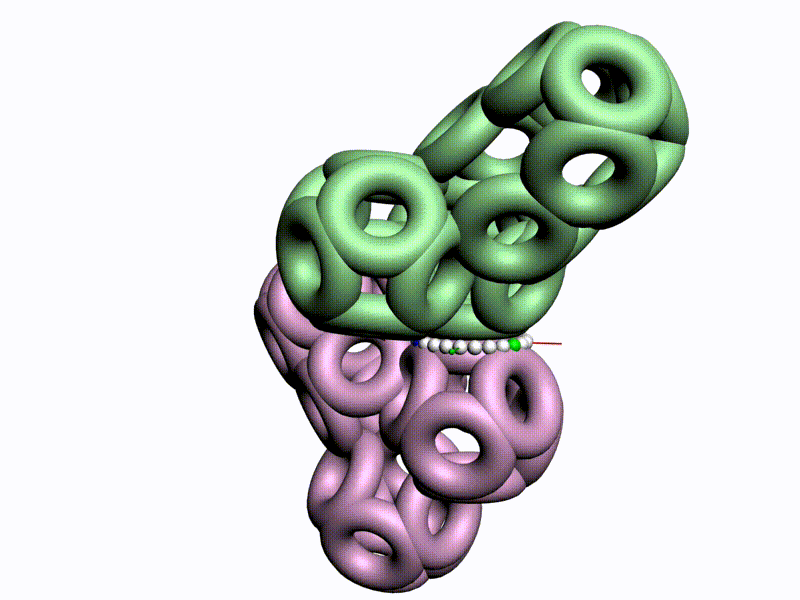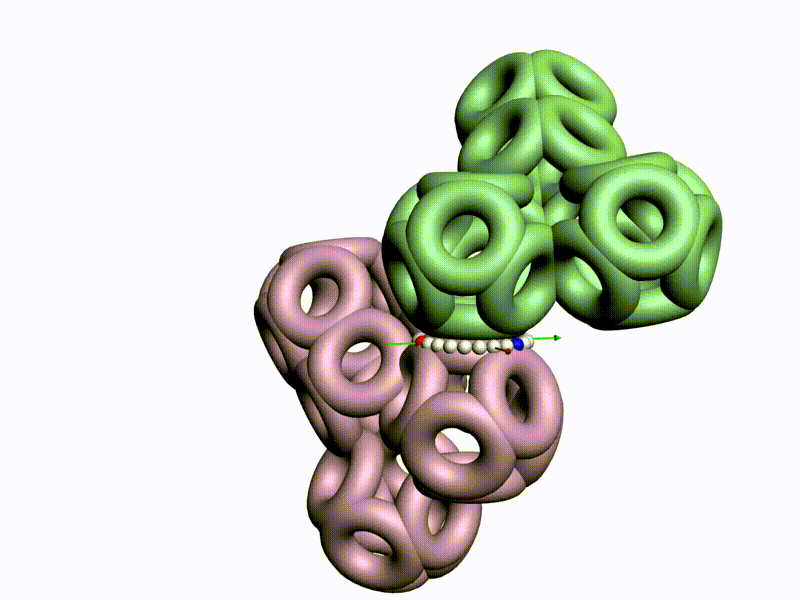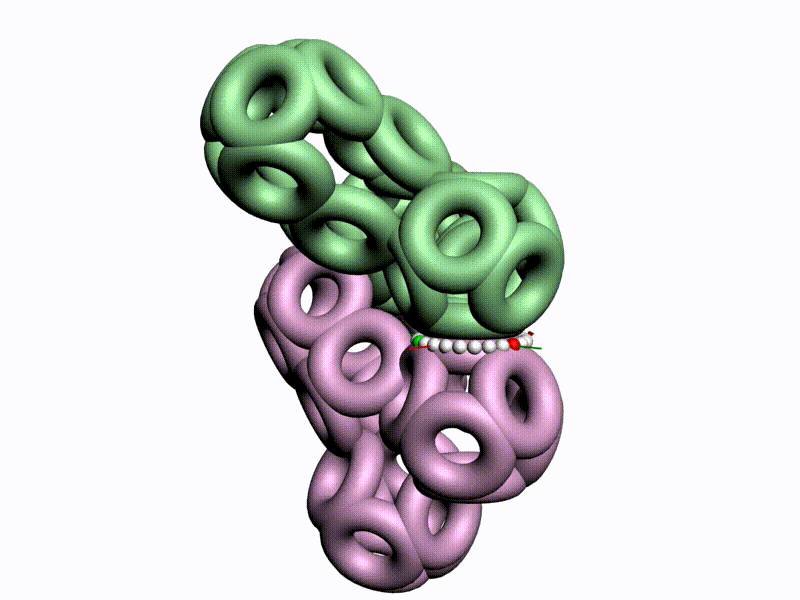

{kind=link}
