| ← Декабрь 2014 → | ||||||
|
1
|
2
|
4
|
||||
|
8
|
9
|
12
|
13
|
14
|
||
|
16
|
17
|
19
|
21
|
|||
|
22
|
24
|
28
|
||||
|
30
|
31
|
|||||
За последние 60 дней 20 выпусков (2-3 раза в неделю)
Сайт рассылки:
http://nanoworld.narod.ru
Открыта:
07-10-2005
Статистика
-1 за неделю
415 Опытное производство для шлифовки рубиновых шариков
|
Выпуск 415 Лаборатория Наномир Когда реальность открывает тайны, уходят в тень и меркнут чудеса ... Опытное производство для шлифовки рубиновых шариков Приблизительно так будет выглядеть процесс шлифовки рубиновых шариков: Видео: https://yadi.sk/i/Kh48OaVZcjLT3 Skype: [2:05:11] s: Я со всеми вашими устройствами не знаком. Можете
рассказать суть, а так можно подумать о покупке или
дофинансировании [10:07:56] Кушелев Александр Юрьевич: В г.Орле заканчивают
делать модулятор для 30-киловаттного магнетрона МИ-88 [10:08:25] Кушелев
Александр Юрьевич: От него мы планируем включить рубиновый источник энергии,
состоящий из 3 рубиновых шариков диаметром 8.3 мм [10:09:17] Кушелев
Александр Юрьевич: Далее, через переходные размеры 8.2, 8.1 планируется включить
десятка полтора рубиновых генераторов, состоящих из шариков диаметрами 8.0
мм [10:11:05] Кушелев Александр Юрьевич: Но для промышленной партии нужно
заказать 10 000 рубиновых шариков диаметром 5.0 мм и часть номинального ряда
8...5 мм, чтобы включить 2200 генераторов 5-мм диапазона. Из них будут
собираться и летательные браслеты. Комплект из 8 и 12 браслетов сможет дать
суммарную силу до 200 и 300 кГ. [10:12:40] Кушелев Александр Юрьевич:
Подробнее о создании рубиновой энергетики можно прочесть в рассылке и на форуме
лаборатории Наномир: http://nanoworld.org.ru/topic/760/ [10:13:10]
Кушелев Александр Юрьевич: http://nanoworld.org.ru/topic/727/ [10:13:29]
Кушелев Александр Юрьевич: http://nanoworld.org.ru/topic/894/ [10:13:58]
Кушелев Александр Юрьевич: Рассылка: http://subscribe.ru/catalog/science.news.nanoworldnews [10:14:41] Кушелев Александр Юрьевич: Копия рассылки с вырезанной рекламой и восстановленными картинками: http://nanoworld88.narod.ru/data/index.htm [17:05:40] Кушелев Александр Юрьевич: Процесс шлифовки рубиновых шарков на станке стартовал: https://yadi.sk/i/L26QatAncjtXP
[17:05:58] Кушелев Александр Юрьевич (инвестору С): Благодаря Вам удалось заказать стёкла для шлифовки шариков на станке [17:15:07] Инвестор С: здорово) [17:15:14] Инвестор С: есть
какие-то
результаты? [17:16:26] Кушелев Александр Юрьевич: Станок запущен, шарики 8.05
... 8.03 начали дорабатываться. Через неделю, вероятно, доработаются [17:17:01] Кушелев Александр Юрьевич: Уже хорошо. Вручную-то я уже
замучился. Сделал 15 генераторов и устал. [17:17:20] Инвестор С: конечно
тяжко.. [17:17:27] Кушелев Александр Юрьевич: А станок может круглосуточно
работать [17:17:33] Кушелев Александр Юрьевич: Потребление 20
Вт [17:17:48] Кушелев Александр Юрьевич: Шарики катаются между алмазной
планшайбой и стеклом [17:18:41] Кушелев Александр Юрьевич: А 5-мм шарики
китайцы могут сделать сразу с микронной точностью. У них там на этот размер
оборудование, вероятно, хорошее [17:18:46] Инвестор С: главное первые
получить и посмотреть результат [17:19:09] Кушелев Александр Юрьевич:
Главное, чтобы этот результат никто не спрятал на много лет [17:19:10]
Инвестор С: они там еще не заподозрили ничего?) [17:19:29] Кушелев Александр
Юрьевич: Китайцы рубиновые шарики миллиардами штук в год выпускают [17:19:35]
Кушелев Александр Юрьевич: Это всё идёт на ювелирку [17:19:42] Кушелев
Александр Юрьевич: шарики гранят и делают бусы [17:19:53] Кушелев Александр
Юрьевич: Но нам нужны пока шарики [17:20:02] Кушелев Александр Юрьевич:
Гранить будем позже и точнее [17:20:10] Инвестор С: для др
моделей? [17:20:14] Кушелев Александр Юрьевич: Да [17:20:27] Инвестор С:
гранить сложнее чем сразу шарик? [17:20:33] Инвестор С: в плане не проще
гранить сразу? [17:20:40] Кушелев Александр Юрьевич: Гранить сложнее, т.к.
нужна высокая угловая точность [17:20:59] Кушелев Александр Юрьевич: Гранить
только с шариков получается [17:21:11] Кушелев Александр Юрьевич: Тем более,
что грани обычно неполные [17:21:28] Инвестор С: там выше Q? [17:21:32]
Кушелев Александр Юрьевич: А можно на шариках резьбу нарезать, но китайцы пока
не берутся [17:21:34] Инвестор С: при гранях [17:22:01] Кушелев Александр
Юрьевич: У гладких шариков максимальная добротность, если 10 или больше длин
волн по экватору [17:22:16] Кушелев Александр Юрьевич: А в гранёных можно
уменьшить число волн, но точность нужна высокая [17:22:35] Кушелев Александр
Юрьевич: Может быть галтовка позволит снизить требования к точности
огранки [17:22:44] Кушелев Александр Юрьевич: Но тут нужно ещё
экспериментировать [17:23:06] Инвестор С: т.е. понизить частоту для граненых,
но что это даст в плане W (энергии)? [17:23:15] Кушелев
Александр Юрьевич: Я пока получил добротность галтованных кубиков (не очень
точно огранёных) около 20 000, а нужно 30 000 и выше [17:23:35] Кушелев
Александр Юрьевич: Гранёные позволяют уменьшить количество
материала [17:23:46] Инвестор С: разумно да [17:23:46] Кушелев Александр
Юрьевич: Чем меньше экватор, тем меньше объём шарика [17:23:54] Инвестор С:
но в принципе это не важно) [17:23:55] Кушелев Александр Юрьевич: Я уже
кубики проверял [17:23:58] Инвестор С: при таких ценах  В
качестве дополнительного груза используется сковородка  Станок за 8 часов уменьшил асферичность самых крупных
шариков с 20 до 4 микрон. Ещё сутки, и первая партия (94 шарика), вероятно,
будет готова.  Смещение
стекла приводит к равномерному снятию материала по
всей площади.  Процесс
шлифовки идёт правильно, но ... абразив практически
закончился. Без основной части (1000 карат) дошлифовать шарики не удастся.
Первая партия рубиновых шариков из Китая доработана. Все шарики, которые имели размер больше 8.035, стали иметь диаметр 8.035 мм с асферичностью меньше микрона. Те, что были меньше, ещё предстоит дошлифовать...
Инвесторы планируют заказать 10 000 рубиновых шариков диаметром 5 мм. Эти шарики у китайцев особенно точно получаются. Из 10 000 шариков можно будет собрать 2000 источников энергии по 5 кВт.
Первый рубиновый генератор, собранный из доработанных китайских шариков.  Первые 30 рубиновых генераторов для инвесторов.
Это упакованные для отправки на эксперимент 98 рубиновых генераторов для инвесторов:
[17:24:03] Кушелев Александр Юрьевич: Но шарики - не самый крутой
вариант. Круче яйца [17:24:12] Кушелев Александр Юрьевич: В яйце всего две пучности волны [17:24:21] Инвестор С: яйцо - ОСНОВА жизи) [17:24:34] Кушелев Александр Юрьевич: Но непонятно пока, как делать точные яйца [17:24:36] Инвестор С: а там наверно должна быть
формула ПРАВИЛЬНОГО яйца еще.. [17:24:43] Инвестор С: вово [17:24:46] Кушелев Александр Юрьевич: Я уже в рассылке опубликовал [17:25:04] Кушелев Александр Юрьевич: Инопланетяне не заморачиваются с точностью морских камней в форме яйца [17:25:11] Кушелев Александр
Юрьевич: Они просто их юстируют [17:25:14] Кушелев Александр Юрьевич: Сейчас покажу [17:25:21] Инвестор С: подбирают f? [17:26:06] Кушелев Александр Юрьевич: На фотографии видны насечки на каменном яйце [17:26:11] Кушелев Александр Юрьевич: Это и есть результат
юстировки [17:26:24] Кушелев Александр Юрьевич: После юстировки кривоватое яйцо имеет идеальное поле [17:27:16] Инвестор С: причем не все линии строго перпендикулярны [17:27:40] Инвестор С: на фото камень вроде не 2.5 метра - трава высокая.. [17:29:06] Кушелев Александр Юрьевич: Это другой камень. Фото яйца у истока озера Рица мне не удалось достать. Мне о нём местный абхазец рассказывал. Он его сфотографировал на мобильник, но фотка не сохранилась. [17:35:25] Инвестор С: а фото этого из какого места? [17:35:50] Кушелев
Александр Юрьевич: Это, вероятно, из Англии [17:37:28] Инвестор С: получается
ОНИ его облучают, смотрят дисбаланс как на "шиномонтаже", потом его корректируют
надрезами? [17:37:43] Кушелев Александр Юрьевич: Да [17:37:48] Кушелев
Александр Юрьевич: Я тоже так могу [17:37:59] Инвестор С: по добротности? [17:38:03] Кушелев Александр Юрьевич: Только сразу не догадаться,
что камень с надрезами - резонатор [17:38:17] Инвестор С: по пику
Q? [17:38:19] Кушелев Александр Юрьевич: Добротность при таких размерах у
гранита может быть за 100 000 [17:38:37] Инвестор С: кстати у многих пирамид
- прослойка слюды [17:38:49] Инвестор С: наверно тоже для каких-то подобных
целей [17:39:56] Кушелев Александр Юрьевич:
[17:40:12] Кушелев Александр Юрьевич: Это я начинал моделирование: http://nanoworld88.narod.ru/data/171.htm [17:41:10]
Кушелев Александр Юрьевич:
[17:41:24] Кушелев Александр Юрьевич: Продолжение: http://nanoworld88.narod.ru/data/173.htm [17:42:25] Кушелев Александр Юрьевич: А здесь уже по полной программе: http://nanoworld88.narod.ru/data/174.htm [17:43:01]
Кушелев Александр Юрьевич: Оказалось, что "сердце" - это разрезанное по
диагонали "яйцо" и половинка повёрнута. [17:43:57] Кушелев Александр Юрьевич:
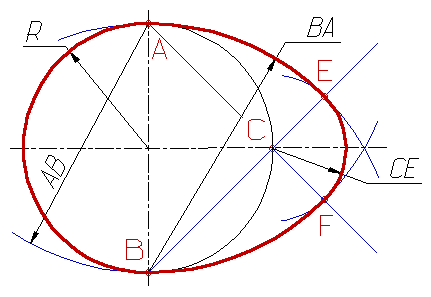
В том же 174-ом выпуске рассылки есть и моделирование яйца. Африканский крест, вписанный в ... яйцо?! Здесь интересны не только бусы, но и необычый крестик на лбу...
Из 4 одинаковых элементов резонансной системы изменены две. Причём увеличены в разной степени. На первом этапе крест в круге превращается в вытянутый крст в эллипсе или овале. На втором этапе овал превращается в яйцис (сечение яйца вдоль большой полуоси). На каждом этапе преобразования формы мощность резонансной системы увеличивается. Кроме того, после превращения овала (эллипса) в яйцис, источник энергии превращается в двигатель. В этом плане интересно прследить, как двигатель-крестояйцис превратится в фигуру вращения, т.е. в более мощный двигатель-яйцо.
Мобильный измерительный СВЧ-генератор Материал с форума лаборатории Наномир: diprospan: Кушелев: Огромное спасибо! Это как раз то, что нужно для исследования резонансных свойств мегалитических комплексов и их элементов! Вам всегда удаётся найти очень важную для лаборатории Наномир информацию.
Сейчас попробую заказать этот гаджет, который позволит донести измерительную
аппаратуру в горы без верблюдов и лошадей Чего не хватает для светлого будущего человечества? Skype, 2014-11-19:
[4:23:39] Кушелев Александр Юрьевич: Длительность импульса 0.2
мкс - в 10 раз меньше, чем делает Золдракс. Но начать можно и с такого. А вдруг
хватит? [4:28:42] Кушелев Александр Юрьевич: Магнетрон может дать
0.6 мкс. Это уже лучше. [4:29:10] Кушелев Александр Юрьевич: Модулятор может
дать 0.5 мкс [4:29:40] Кушелев Александр Юрьевич: Золдракс делает 2.0
мкс [4:31:37] Кушелев Александр Юрьевич: 2 мкс ~ 70 000 периодов несущей. Со
стабилизацией амплитуды должно хватить. [4:32:31] Кушелев Александр Юрьевич:
А какова форма импульса в модуляторе с Тантала? Если не стабилизированный
импульс, то скорее всего не подойдёт. [4:32:56] Кушелев Александр Юрьевич: А стабилизатор как раз Золдракс сейчас и делает.
Сапоги-скороходы, точнее скоролёты с орнаментом источника энергии, включающим такие элементы, как свастика...
Источник энергии имеет вид барельефной свастики, расположенной в диэлектрическом барельефном экране.
Так будут выглядеть барельефные орнаменты источников энергии во включенном состоянии:
Измерительная рулетка Кришны "Кришна ворует масло", - так называется эта картина. Обратите внимание на чередование цветных бусин. Мы видим изображение точной рулетки с цветовым кодом. Марья-искусница вышивает QR-код лаборатории Наномир
Пока один станок шлифует китайские рубиновые шарики, я собрал ещё один станок для изготовления источника энергии с QR-кодом лаборатории Наномир.
Видео: https://yadi.sk/i/D6sJx4a4cmLu7 Пока делаю модель из стеклянных шариков, т.к. сапфировых и рубиновых в нужном количестве ещё нет. Нужно заказывать в Китае. На этом светлые бусинки закончились. Чуть-чуть не хватило.
Модель рубинового/сапфирового источника энергии с QR-кодом лаборатори Наномир удалось закончить с ипользованием бусин других цветов.
QR-код распознаётся декодером: http://zxing.org/w/decode.jspx
QR-код распознаётся декодером: http://zxing.org/w/decode.jspx
На таком фоне декодер из инета не может распознать QR-код, но гаджеты типа айфона или андроида смогут.
А в таком варианте QR-код не распознает ни один гаджет Кодирование вторичной структуры и структурного шаблона белка в геноме эукариот Соколик Виктория Васильевна Виктория Соколик: На III международной научно-практической конференции: «Современная биология: актуальные вопросы» 14-15.11.2014 город Санкт-Петербург, Россия была опубликована моя статья по пикотехнологии. Таким нехитрым способом пытаюсь достучаться до сознания ученых в этой области. Всем фанатам Александра Юрьевича настоятельно рекомендую присмотреться к списку литературы, в котором я ссылаюсь и на его раннюю работу (№4) и на нашу совместную публикацию (№14))). КОДИРОВАНИЕ ВТОРИЧНОЙ СТРУКТУРЫ И СТРУКТУРНОГО ШАБЛОНА БЕЛКА В ГЕНОМЕ ЭУКАРИОТ Соколик Виктория Васильевна кандидат биологических наук, ведущий научный сотрудник ГУ «Институт неврологии, психиатрии и наркологии НАМН Украины», г. Харьков ENCODING OF SECOND STRUCTURE AND STRUCTURAL TEMPLATE OF PROTEIN IN EUKARIOTIC GENOME Sokolik Victoria Candidate of Science, principal research fellow SI “Institute of neurology, psychiatry and narcology of the NAMS Ukraine” Kharkov АННОТАЦИЯ Цель. Обосновать кодирование вторичной структуры и её топологии в структурном шаблоне белка в геноме эукариот. Методы. Анализировали вторичную структуру 100 белков эукариот (PDB). Оценивали частоту встречаемости кодонов XYC/G, XYА, XYТ и их кластеров в нуклеотидных последовательностях, детерминирующих спирали, β-тяжи, повороты и неструктурированные фрагменты белков. Результаты обрабатывали статистически. Результаты. Установили, что правая спираль кодируется (XYC/G)n, n3, повороты – (XYA)n, n3, β-тяжи – XYT. Вывод. В геноме эукариот содержится информация о структуре белков. ABSTRACT Background. Substantiate the encoding of the secondary structure and its topology in the structural template of protein in eukaryotic genome. Methods. Analyzed a secondary structure of 100 eukaryotic proteins (PDB). Evaluated the frequency of codons XYC/G, XYA, XYT and their clusters in nucleotide sequences, which determine the helix, β-strands, turns and unstructured protein fragments. The results were treated statistically. Result. Found that the right helix encoded (XYC/G)n, n3, turns – (XYA) n, n3, β-strands – XYT. Conclusion. In eukaryotic genome contains information about the structure of proteins. Ключевые слова: вторичная структура; структурный шаблон белка; генетический код. Keywords: second structure; structural template of protein; genetic code. В настоящее время одним из алгоритмов моделирования пространственной структуры белка in silico, который исходит из его аминокислотной последовательности, является способ построение модели по гомологии на основе общего структурного шаблона гомологичных белков [1-3]. Десять лет назад Кушелевым А.Ю. была сформулирована идея кодирования пространственной структуры белка [4], которая в абстрактном виде звучала ещё в первых работах Gamow G. [5]. Дальнейшая разработка проблемы привела Соколик В.В. к гипотезе о том, что в ходе трансляции на рибосоме синтезируется не только аминокислотная цепочка, как считали ранее [6-7], и не полностью спирализованный полипептид, как недавно предположил Спирин А.С. [8], а индивидуальный структурный шаблон белка со специфичной топологией вторичной структуры [9-12]. При этом информация третьего нуклеотида детерминирует реализацию элементов вторичной структуры в структурном шаблоне белка (табл. 1) с помощью пула изоакцепторных тРНК для синонимичных кодонов [13]. Механизм такой реализации состоит в декодировании, формировании и фиксировании в полипептидном каркасе синтезируемого белка ротамеров пептидной связи R, L и 0, которые различаются между собой углом поворота θ, кратным 120о (рис. 1). Крайне важно понимать, что ротамер пептидной связи реализуется в процессе матричного синтеза белка в рибосоме и дальнейшее вращение по уже образовавшейся полуторной пептидной связи становиться невозможным. Поэтому с рибосомы сходят совершенно индивидуальные, но, в тоже время, идентичные между собой, молекулы структурных шаблонов синтезируемого белка, состоящие из последовательности ротамеров пептидной связи между аминокислотными остатками, в соответствие с информацией, содержащейся в его гене.
Таким образом, цель исследования состояла в обосновании кодирования в геноме вторичной структуры и её топологии в структурном шаблоне белка. Дизайн исследования. Проанализировали случайную выборку из 100 белков эукариот, для которых в базе данных PDB представлены схемы вторичной структуры. Оценивали абсолютную и относительную частоту встречаемости кодонов вида XYC/G, XYА, XYТ и их кластеров (XYC/G)n, (XYА)n, (XYТ)n, n3, в нуклеотидных последовательностях, детерминирующих спирали, β-тяжи, повороты и неструктурированные мотивы белков. Результаты обрабатывали статистически: t-критерий Стьюдента использовали для анализа производных показателей (абсолютная и относительная частота), убедившись в нормальности распределения; P-тест Фишера и четырехпольные таблицы – для выявления статистической зависимости между конформерами и разновидностями кодонов и/или их кластеров: χ2 ˃ χ2крит (3,8). Различие считали статистически значимым при p 0,05, тенденция к значимым отличиям при p 0,10. Результаты. Установили, что правая спираль кодируется кластерами кодонов (XYC/G)n, n3 (Р = 7.33·10-22, χ 2 = 42.62). Повороты детерминированы кластерами кодонов (XYA)n, n3 (Р = 2.18 · 10-3, χ2 = 6.95). Эти данные согласуются с выявленным достоверным превышением относительной частоты встречаемости кластеров (XYC/G)n, n3 и (XYA)n, n3 в областях генов, кодирующих спиральные фрагменты и повороты, соответственно (рис. 2). β-Тяжи менее жестко детерминированы кодонами XYT. Тест Фишера показал кодирование β-тяжей кодонами XYT или их кластерами (XYТ)n, n3: Р = 3.63·10-6 (χ 2 = 22.05) и Р = 3.43·10-2 (χ 2 = 4.56), соответственно, для изучаемого пула белков. Для конформеров вторичной структуры белка характерна периодичность. Поэтому правая спираль в качестве структурного шаблона для правой α-, правой π- или правой 3/10 спиралей в белке кодируется последовательностью кодонов с С/G в третьей позиции (n 3), β-спираль (прототип β-тяжа) – кодонами с Т, а левая спираль (или её единичный виток – прототип поворота) – последовательностью кодонов с А в третьем положении триплета (n 3). Неструктурированные фрагменты белка кодируются чередованием кодонов с С, G, А и Т в третьей позиции кодонов, что обусловливает отсутствие периодичности [14].
Таблица 1 легла в основу алгоритма авторской программы Молекулярный конструктор (МС), которая декодирует нуклеотидную последовательность с созданием координатного файла и визуализацией индивидуального структурного шаблона белка в качестве основы для дальнейших исследований методами молекулярной динамики. Данный механизм кодирования и трансляции генетической информации является эволюционно новым. Его формирование у эукариот было обусловлено необходимостью синтеза больших и сложных белков в виде структурных шаблонов, максимально приближенных к функциональным конформациям этих белков, чтобы фолдинг имел наибольшие скорость и КПД. У прокариот и органелл эукариот (митохондрии, хлоропласты) третий нуклеотид кодонов в генах небольших полипептидов ещё не является информационным, поэтому на нём и наблюдается воблирование по описанному Crick F. механизму [15]. Это обусловлено отсутствием пула изоакцепторных тРНК со специфически модифицированными нуклеотидами в первом положении антикодона и нередко отсутствием филогенетически более молодых областей в структуре тРНК [16], т.е. недоразвитием звена, реализующего информацию третьего нуклеотида. Анализ пространственной структуры семейств изоакцепторных тРНК с синонимичными кодонами позволил автору предложить гипотезу о перекодировании информации третьего нуклеотида кодонов в соответствующий ротамер пептидной связи непосредственно самой 3D-структурой тРНК, а точнее углом поворота акцепторного стебля вокруг своей оси. Модификации первого нуклеотида антикодонов тРНК лишь способствуют точности считывания этой информации и однозначности взаимодействия с мРНК в рибосоме [13-14]. В последнее время догма о несущественной роли третьего нуклеотида кодонов для синтеза белка была опровергнута результатами экспериментов по синонимическому мутагенезу. В частности было показано, что даже единственная синонимическая замена кодона может иметь значительное влияние на уровень экспрессии генов, фолдинг белков и на пространственную структуру последних [17-18]. Поэтому синонимичные кодоны (отличающиеся нуклеотидом в третьем положении триплета, но кодирующие одну и ту же аминокислоту) не являются взаимозаменяемыми, даже если они неразличимы в отношении аминокислот [19]. Всё чаще высказывается мнение, что избыточность генетического кода сформировалась как способ сохранения структурной информации о белках в кодирующих их нуклеотидных последовательностях [20]. Однако боль-шинство исследователей отмечают, прежде всего, связь частоты использования синонимичных кодонов со скоростью процесса матричного синтеза белка, полагая, что низкочастотные «медленные» кодоны в кодирующей нуклеотидной последовательности обеспечивают генетическую инструкцию, которая регулирует скорость синтеза белка, чтобы позволить растущей полипептидной цепи сформировать вторичную и третичную структуры [20-23]. Результаты данной работы, напротив, демонстрируют наличие в генах белков не только временных, а и пространственных инструкций для однозначного матричного синтеза уникальных структурных шаблонов белков. Вывод. В геноме эукариот содержится информация о структуре белков. Список литературы 1. Zhang Y. 2008. Progress and challenges in protein structure prediction. Curr Opin Struct Biol 18 (3): 342–348. 2. Nayeem A., Sitkoff D., Krystek S. Jr. 2006. A comparative study of available software for high-accuracy homology modeling: From sequence alignments to structural models. Protein Sci 15: 808–824. 3. Sivasubramanian A., Sircar A., Chaudhury S., Gray J. J. 2009. Toward high-resolution homology modeling of antibody Fv regions and application to antibody–antigen docking. Proteins 74: 497–514. 4. Кушелев А.Ю., Полищук С.Е., Неделько Е.В. 2002. Построение масштабной модели структуры белка. Актуальные проблемы современной науки. 2: С. 236–243. 5. Gamow G. 1954. Possible Relation between Deoxyribonucleic Acid and Protein Structures. Nature 173: 318. 6. Crick F.H., Barnett L., Brenner S., Watts-Tobin R.J. 1961. General nature of the genetic code for proteins. Nature. 192: 1227-1232. 7. Азимов А. 2006. Генетический код. От теории эволюции до расшифровки ДНК. М.: Центрполиграф: 300. 8. Спирин А.С. 2011. Молекулярная биология: рибосомы и биосинтез белка. М.: «Академия»: 496. 9. Sokolik V.V. 2011. Protein is coded in genome and synthesized in ribosomes as a structural template of a rotameric version sequence of peptide bound configuration. МССМВ-11: 347–348. 10. Sokolik V.V. 2010. Modeling of the individual structural template of protein on determining it nucleotide sequences. BGRS\SB-2010: 275. 11. Соколик В.В. 2012. Предсказание пространственной структуры белка in silico на основе информации генома и геометрического алгоритма – альтернатива квантово-механическому подходу. Материалы Международной научной конференции «Математическое и компьютерное моделирование в биологии и химии. Перспективы развития»: С.155-158. 12. Соколик В.В. 2014. Никакой дополнительной информации, большей, чем та, что содержится в ДНК, для сворачивания белка не требуется. Ukr. Biochem. J. 86(5), Suppl. 1: 37-38. 13. Соколик В.В. 2011. Загадка изоакцепторных тРНК. II Всероссийская Интернет-Конф. «Актуальные проблемы биохимии и бионанотехнологии»: 11-15. 14. Кушелев А.Ю., Соколик В.В. 2012. Пикотехнология – новый подход в моделировании пространственной структуры белка. Заочная Междунар. научно-практическая конференция «Современная наука: тенденции развития»: 203-207. 15. Crick F.H.C. 1968. The Origin of the Genetic Code. J. Mol. Biol. 38: 367–379. 16. Maizels N., Weiner A.M. 1994. Phylogeny from function: Evidence from the molecular fossil record that tRNA originated in replication, not translation. Proc. Nat. Acad. Sci. USA. 91(15): 6729–6734. 17. Saunders R., Deane Ch.M. 2010. Synonymous codon usage influences the local protein structure observed. Nucleic Acids Res. 38: 6719–6728. 18. Plotkin J.B., Kudla G. 2011. Synonymous but not the same: the causes and consequences of codon bias. Nat. Rev. Genet. 12: 32–42. 19. Biro J.C. 2008. Correlation between nucleotide composition and folding energy of coding sequences with special attention to wobble bases. Theor. Biol. Med. Model. 5: 14. 20. Angov E. 2011. Codon usage: Nature's roadmap to expression and folding of proteins. Biotechnol J. 6: 650–659. 21. Marin M. 2008. Folding at the rhythm of the rare codon beat. Biotechnol. J. 3: 1047–1057. 22. Spencer P.S., Barral J.M. 2012. Genetic code redundancy and its influence on the encoded polypeptides. Computational and Structural Biotechnology J. 1: 1–8. 23. Grosjean H., de Crecy-Lagard V., Marck C. 2010. Deciphering synonymous codons in the three domains of life: co-evolution with specific tRNA modification enzymes. FEBS Lett. 584: 252–264.
|






































| В избранное | ||
